KANG Jianjun, ZHAO Ming, TAN Yanrong, ZHU li, BING Danhui, ZHANG Yangdong, TONG Shengli. , Sand-fixing characteristics of Carex brunnescens and its application with straw checkerboard technique in restoration of degraded alpine meadows . Journal of Arid Land, 2017, 9(5): 651-665.
Sand-fixing characteristics of Carex brunnescens and its application with straw checkerboard technique in restoration of degraded alpine meadows
KANG Jianjun1,2, ZHAO Ming1,*, TAN Yanrong1, ZHU li1, BING Danhui1, ZHANG Yangdong1, TONG Shengli3
1Gansu Research Academy of Forestry Science and Technology, Lanzhou 730000, China
2Northwest Institute of Eco-Environment and Resources, Chinese Academy of Sciences, Lanzhou 730000, China
3Administration of Gansu Xinglongshan National Nature Reserve, Lanzhou 730000, China
*Corresponding author: ZHAO Ming (E-mail: msdse9@126.com)
Abstract
Carex brunnescens (Pers.), a typical clonal species, is considered to be the only herb found to date that can develop on sand dunes in Maqu alpine region of northwestern China. However, the characteristics that C. brunnescens resists to harsh alpine environment have not been documented. In this study, we conducted a field investigation to determine the morphological, reproductive, and sand-fixing characteristics of C. brunnescens. Concomitantly, we transplanted the belowground rhizomes of C. brunnescens to sand dunes and compared the abilities to restore degraded alpine meadows among sand dunes that had no further treatment (SD+N), sand dunes that had straw checkerboard technique but no transplanted rhizomes of C. brunnescens (SD+SCT), and sand dunes that had both SCT and transplanted rhizomes of C. brunnescens (SD+SCT+P). We found that belowground vertical rhizomes and horizontal rhizomes (including branching rhizomes and main rhizomes) of C. brunnescens were highly developed and that population reproduction was dominated by horizontal rhizomes. C. brunnescens exhibited a significant sand-fixation effect under following conditions: population density was 145-156 ramets/m2, vegetation cover was 31.2%-39.3%, total length of belowground rhizomes was 11,223 cm/m2, total length of belowground first-order roots was 9161-10,524 cm/m2, fresh weight of aboveground part was 198.5-212.6 g/m2, and fresh weight of belowground part was 578.8-612.4 g/m2. It should be particularly noted that SD+SCT+P treatment (sand dunes that had both straw checkerboard technique and transplanted rhizomes of C. brunnescens) was the best and SD+N (sand dunes that had no further treatment) was the worst in terms of following biotic indicators: total number of reproductive ramets, total number of belowground rhizomes, and fresh weight of aboveground and belowground parts of C. brunnescens, contents of soil organic carbon, available nitrogen, microbial biomass carbon, and microbial biomass nitrogen. It implies that applying SCT in sand dunes and transplanting belowground rhizomes to sand dunes with SCT could improve both soil fertility and growth of C. brunnescens. These results suggest that the SCT-promoted high reproductive abilities of belowground rhizomes of C. brunnescens can successfully facilitate the establishment of ramets and can thus be an effective strategy to restore degraded vegetation in Maqu alpine region of northwestern China.
Desertification is a process of land degradation resulting from climatic changes and human activities in arid and semi-arid areas. Desertification involves severe vegetation degradation and intensified wind and water erosions (Zhao et al., 2009). In particular, desertification caused by wind erosion poses a severe threat to agricultural productivity and ecological environment (Gomes et al., 2003; Lian et al., 2013). Even worse, regional political instability and social insecurity have also been linked to desertification (Meadows and Hoffman, 2002).
Sand burial and wind erosion are two of the most serious abiotic stresses on plant growth in many arid and semi-arid areas (Li et al., 2010a; Luo and Zhao, 2015). Deep sand burial can significantly inhibit plant growth and succession. However, stimulating effects on plant growth were also observed when plants were subjected to relatively shallow sand burial (Dong et al., 2000; Dech and Maun, 2006; Zhao et al., 2007; Jiang et al., 2016). Wind erosion can render soil infertile through the removal of nutrient-rich surface soil and a progressive decrease in vegetation cover (Yu et al., 2008; Li et al., 2010a, b). Clonal plants, the dominant plant type growing in sand dune environments, have developed multiple adaptive strategies to resist sand burial and wind erosion (Roiloa et al., 2014; Luo and Zhao, 2015). Clonal plants can generate new offspring ramets connected with parent plants. Relative to parent plants, the new ramets could achieve clonal expansion horizontally and thus enter into different habitats (Ye and Dong, 2011; Kang et al., 2016a). Clonal integration is considered to be one of the most important strategies for the survival of clonal plants because it allows buried ramets to access to water and nutrients via non-buried ramets, thus ensuring plant survival and reproduction in the environments that are prone to sand burial and wind erosion (Amsberry et al., 2000; Xu et al., 2010; Luo et al., 2015).
Maqu is one of the most important alpine meadows at the eastern edge of the Tibetan Plateau in northwestern China. The area is known as “ the kidney of the Yellow River” because of its vital function in conserving soil and water in the upper reaches of the Yellow River (Kang et al., 2015). However, long-term over-exploitation of alpine meadow associated with rapid population increases, environmentally-unfriendly means of agricultural production, overgrazing by livestock, mining, and urbanization inevitably triggered severe desertification in this region (Qi et al., 2006; Wei et al., 2010). The situation has been worsened by human alteration of river channels and also by human withdrawal of groundwater in the Yellow River (Wang et al., 2001, 2005). Consequently, the Maqu alpine region has risk to become a main source of sandstorms in China if effective control measures are not implemented (Wang et al., 2008).
Carex brunnescens(Pers.) is considered to be the only herb species found to date that can develop on sand dunes in Maqu alpine region (Kang et al., 2016a, b; Ma et al., 2017). This species is a typical clonal plant that is reproduces by forming strong belowground horizontal rhizomes. Due to low seed germination rate, C. brunnescens rarely reproduces sexually even in high-precipitation months or in wet years (Wang et al., 2013; Zhu et al., 2013). C. brunnescens is highly tolerant to wind erosion and also to sand burial because of its rapid growth and regeneration capacities of rhizomes, basal meristem and belowground storage organs (Kang et al., 2016a, b). Recent studies showed that C. brunnescens population can quickly invade and multiply in a new sand habitat with asexual reproduction by belowground rhizomes and thus can stabilize sand dunes. In favored topographical conditions, it can even form rare “ herb dunes” (Kang et al., 2016a, b; Ma et al., 2017). However, studies focusing on how the C. brunnescens resists sand burial and wind erosion are relatively scarce. Further, although the application of C. brunnescens in stabilizing sand dunes is of great interest, the feasibility of combining specific herbaceous plants with other means for enhancing wind protection and sand fixation effect has not been explored. The straw checkerboard technique (SCT) was devised at the Shapotou Desert Research Station, Chinese Academy of Sciences in 1957 (SDRS, 1986) and has been widely used since for stabilizing sand dunes in arid and semi-arid regions of China (Qiu et al., 2004; Ma et al., 2015). SCT refers to a simple mean that straws of wheat, or rice, or reed, or others were placed (with half buried in the sand) in the shape of checkerboard barriers. The straw checkerboard barriers can decrease wind velocity near the ground surface and thus diminish wind erosion of soils. It not only stabilizes the surface of sand dunes but also improves soil qualities through organic inputs of decomposed straw, dead branches and fallen leaves (Jiang et al., 2008; Chen et al., 2013; Kang et al., 2015). In regions where the annual precipitation is very low, some plants such as shrubs of Caragana microphyla Lam., Hedysarum fruticosum Pall. andSalix oritrepha Sohneider and herb of Agropyron cristatum L. can be planted to further improve the windbreak and sand dune fixation effects (Qiu et al., 2004; Jiang et al., 2008; Kang et al., 2013, 2015).
In this study, we selected the typical clonal herbC. brunnescens to determine its morphological, reproductive, and sand-fixing characteristics through a field investigation in a sand dune environment prone to wind erosion and sand burial. Further, we attempted to analyze the influences of belowground rhizomes of C. brunnescens combined with SCT on plant growth, soil fertility, and windbreak and sand dune fixation effects. It is our hope that this study can provide scientific guidelines to explore sand-fixing technology that combines herbs with SCT to recover the degraded vegetation in Maqu alpine region.
2 Materials and methods
2.1 Study area
The study was conducted in about 9.8 km2area with severe wind erosion and sand burial along the banks of the Yellow River in Maqu County, Gansu Province, northwestern China (33° 06′ 30′ ′ -34° 30′ 15′ ′ N, 100° 45′ 45′ ′ -102° 29′ 00′ ′ E; 3700 m a.s.l.). The study area is characterized by a distinct plateau continental climate with annual mean temperature of 1.1° C, mean annual precipitation of 615.5 mm (atmospheric precipitation), and mean annual potential evaporation of 1353.4 mm (Kang et al., 2015). Dominant plant species include herbs such as C. brunnescens and Kobresia robusta Maxim., and shrubs such as S. oritrepha and Hippophae rhamnoides Linn. (Kang et al., 2016b). The landscape consists of fixed, semi-fixed, and moving sand dunes and C. brunnescensvegetation coverage is > 45%, 15%-30%, and < 15%, respectively (Xu and Liu, 2012; Zhang et al., 2016).
2.2 Experimental design
2.2.1 Morphological characteristics of belowground rhizomes of C. brunnescens
In April to September of 2011, we used the randomized complete block design to establish five representative plots (10 m× 10 m each) in the middle parts of the longitudinal semi-fixed and moving sand dunes to determine the morphological characteristics of belowground rhizomes of C. brunnescens. We examined the maximum soil depth of vertical rhizome growth during the vigorous growth period of C. brunnescens in July 2011. Our examination shows that 50 cm is the maximum depth of sand burial for vertical rhizomes of C. brunnescens to grow upwards and to form new ramets. Subsequently, we excavated the belowground rhizomes (to a soil depth of 50 cm) in each plot with three replicates, removed the attached soil by hand shaking, and classified the rhizomes into vertical rhizomes and horizontal rhizomes. We further distinguished two sub-types within the horizontal rhizomes: branching rhizomes and main rhizomes. It should be pointed out that we were not able to completely excavate all horizontal rhizomes in each plot due to the intricacy of the rhizomes. Finally, we measured the length and internode length of the collected vertical rhizomes and horizontal rhizomes (including branching rhizomes and main rhizomes) and counted the number of rhizomes.
2.2.2 Reproductive characteristics of belowground rhizomes of C. brunnescens
Reproductive characteristics of belowground rhizomes of C. brunnescens were observed in a manner similar to that used for the determination of morphological characteristics. Specifically, we used the randomized complete block design to establish four representative plots (5 m× 5 m each) in longitudinal semi-fixed sand dunes to determine the reproductive characteristics of belowground rhizomes of C. brunnescens. We firstly examined the maximum soil depth that belowground rhizomes could survive (0-200 cm). Our examination shows that 120 cm is the maximum depth that belowground rhizomes of C. brunnescens can reach. Then, we divided soil depth of 0-120 cm into 3-4 layers based on the average length (30-40 cm) of vertical rhizomes and classified the rhizomes into vertical rhizomes and horizontal rhizomes (including branching rhizomes and main rhizomes) for each layer. Finally, we observed the reproductive characteristics of belowground rhizomes of C. brunnescens based on the growth performance (including length of rhizomes, internode length of rhizomes, and bud number of rhizome nodes) of vertical and horizontal rhizomes.
2.2.3 Relationships of vegetation parameters with wind erosion and sand burial
When wind erosion was more severe in early April 2011, we observed growth performance (length of rhizomes, internode length of rhizomes, and bud number of rhizome nodes) of C. brunnescens in the direction perpendicular to wind direction. Correspondingly, we determined the spread direction ofC. brunnescens populations for each plot. Then, we used the randomized complete block design to establish ten representatives and replicated belt transects (width of 1 m and length of 17 m) along the spread direction ofC. brunnescenspopulations in longitudinal semi-fixed sand dunes. Among which, five belt transects were used to quantitatively investigate the relationships of population density with wind erosion height and sand burial depth, and the other five were used to establish the relationships of vegetation cover with wind erosion height and sand burial depth. These determinations were conducted once a month from April to September, 2011.
Population density was obtained by excavating all plants in each plot (1 m× 1 m) and counting the ramets. Vegetation cover (projected cover; %) was calculated using the digital imagery (Cailson and Ripley, 1997). Then, sampling points were counted and numbered from photographs and geomorphological characteristics and vegetation status were recorded in April and May. The captured vegetation photographs were adjusted to size (with the removal of length and width of up to 2/3 of the original size) by Photoshop 7.0 software to reduce the errors caused by image edge distortion (Diao et al., 2012).
Wind erosion height and sand burial depth were determined for every wind event in the windy season (April and May) by placing soil sample of known mass in a pan-shaped container (wind erosion pan) and later measured by reweighing the samples (Dai et al., 2011).
We used the randomized complete block design to establish five representative belt transects (width of 1 m and length of 15 m) along the spread direction ofC. brunnescenspopulations in longitudinal semi-fixed sand dunes. Then, the relationships of total length of belowground rhizomes, total length of belowground first-order roots, fresh weight of aboveground part and fresh weight of belowground part with wind erosion height and sand burial depth were quantitatively determined for every month from April to September, 2011.
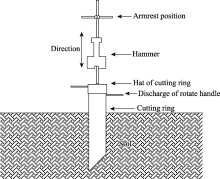
The sampling methods were referred to those described by Kang et al. (2014). Schematic diagram of root sampling instrument is shown in Figure 1. Specifically, along the spread direction ofC. brunnescenspopulations in longitudinal semi-fixed sand dunes, a cutting ring (diameter of 0.15 m and height of 0.5 m) was hammered into the ground vertically allowing for excavation at different soil depths in each quadrat (width of 1 m and length of 1 m) of the belt transect (width of 1 m and length of 17 m), and the ring was then extracted from the soil. The process repeated and transferred to other quadrats. Thus, a large volume of undisturbed soil cores were collected and the characteristics of belowground rhizomes and roots were observed. The aboveground and belowground parts were carefully separated after removing the soil attached to the rhizomes and roots by hand shaking, and their fresh weights were obtained by weighing. Finally, all the belowground rhizomes and roots along the spread direction of C. brunnescens populations were collected to calculate the total length of belowground rhizomes and total length of belowground first-order roots in each quadrat.
Fig. 1 Schematic diagram of root sampling instrument of C. brunnescens plants
2.2.4 Transplanting experiment with the straw checkerboard technique (SCT)
To investigate the effects of SCT on the growth and reproductive performance of belowground rhizomes of C. brunnescens as well as on the soil fertility of C. brunnescens plantations, we conducted a transplanting experiment on sand dunes along the bank of the Yellow River from April 2012 to September 2015. Three experimental sites were designed: sand dunes that had no further treatment (SD+N), sand dunes that had SCT but no transplanted rhizomes of C. brunnescens (SD+SCT), and sand dunes that had both SCT and transplanted rhizomes of C. brunnescens (SD+SCT+P). There were 20 replicated plots in each experimental site. For the straw checkerboard treatments, wheat straw was placed in a 1 m× 1 m checkerboard pattern, and this pattern was reported to be able to achieve remarkable windbreak and sand dune fixation effects (Qiu et al., 2004; Ma et al., 2015). For transplanting of rhizomes of C. brunnescens, healthy and living belowground rhizomes with the same length (25 cm) were collected and placed in a wet plastic bag and then transported to the transplanting sites. We established the transplanting plots in the middle of the longitudinal sand dunes (with application of SCT). In SD+SCT+P experimental site, each transplanting plot (1 m× 1 m; 16 rhizomes per plot covered with 20 cm of wet sand) was surrounded by a buffer zone of 0.5 m. Newly-generated ramets were harvested from each transplanting plot by digging in September 2013, 2014, and 2015. The aboveground and belowground parts were carefully separated and washed three times with water for further morphological analysis.
2.3 Measurements
2.3.1 Growth-related parameters of C. brunnescens
Ramet height was measured with a measuring tape (0.1 m precision). Reproductive ramets and belowground rhizomes were excavated, and the total number was counted in each experimental plot. Total length of belowground rhizomes and total length of belowground first-order roots in each quadrat were calculated by Equation 1. Fresh weight of aboveground part and fresh weight of belowground part in each quadrat were calculated by Equation 2. Survey of plant reproductive characteristics included structural characteristics of rhizomes, source and the maximum sand burial depth from which vertical rhizomes emerged to form new ramets.
where, L (cm) is the total length of belowground rhizomes or total length of belowground first-order roots in each quadrat; L1 (cm) is the total length of belowground rhizomes or total length of belowground first-order roots in each cutting ring; d (m) is the diameter of the cutting ring; h (m) is the height of the cutting ring; W (g) is the fresh weight of aboveground part or fresh weight of belowground part in each quadrat; and W1(g) is the fresh weight of aboveground part or fresh weight of belowground part in each cutting ring, which was obtained by weighing (0.001 g precision).
2.3.2 Soil nutrients
Soil samples were collected in each experimental site (i.e., SD+N, SD+SCT and SD+SCT+P) after all the newly-generated ramets were harvested in September 2015. Specifically, 4-6 subsamples were respectively collected at three soil layers (0-15, 15-30, and 30-50 cm) in each plot, then subsamples from the same soil layer in each plot were mixed into a composite sample. Soil samples were placed in plastic bags and taken to the laboratory for soil chemical analyses. These samples were air-dried at room temperature for 30 d and sieved by a 2-mm mesh. Soil organic carbon (SOC) was measured following the K2Cr2O7-H2SO4 oxidation method of Walkley and Black (Kang et al., 2015), total nitrogen (TN) was measured following the Kjeldahl digestion and total phosphorus (TP) was determined by a UV-1601 visible spectrophotometer (Bao, 2000; Xing et al., 2013). In addition, soil available nitrogen (AN) was determined by the alkaline diffusion method, available phosphorus (AP) was determined by the Bray method, and available potassium (AK) was measured using the ammonium acetate extract method (Bao, 2000).
2.3.3 Soil microbial biomass carbon, nitrogen, and phosphorus
For each experimental site (i.e., SD+N, SD+SCT and SD+SCT+P), five replicated fresh soil samples were collected in each plot. Those soil samples were placed in sealed plastic bags, taken to the laboratory and stored at 4° C. Then, they were fumigated with chloroform after removal of plant residues and other debris. Soil microbial biomass carbon (MBC) was measured by titration with 0.5 mm/L K2SO4 extract, microbial biomass nitrogen (MBN) was determined using the Kjeldahl procedure, and microbial biomass phosphorus (MBP) was measured with the NaHCO3 extraction method after soil fumigation with chloroform (Brookes et al., 1982; Kang et al., 2015).
2.4 Statistical analysis
In this study, we used one-way analysis of variance (ANOVA) for SPSS 13.0 (SPSS Inc., Chicago, IL., USA) to determine the treatments-resulted differences in growth-related parameters of C. brunnescens (such as ramet height, total number of reproductive ramets, fresh weight, etc.) and in soil properties (such as SOC, TN, TP, TK, etc.). Duncan’ s multiple range tests for a completely randomized design were used to detect significant differences in treatment means at P< 0.05 level.
3 Results
3.1 Reproductive characteristics of belowground rhizomes of C. brunnescens
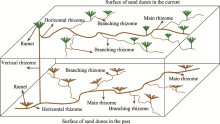
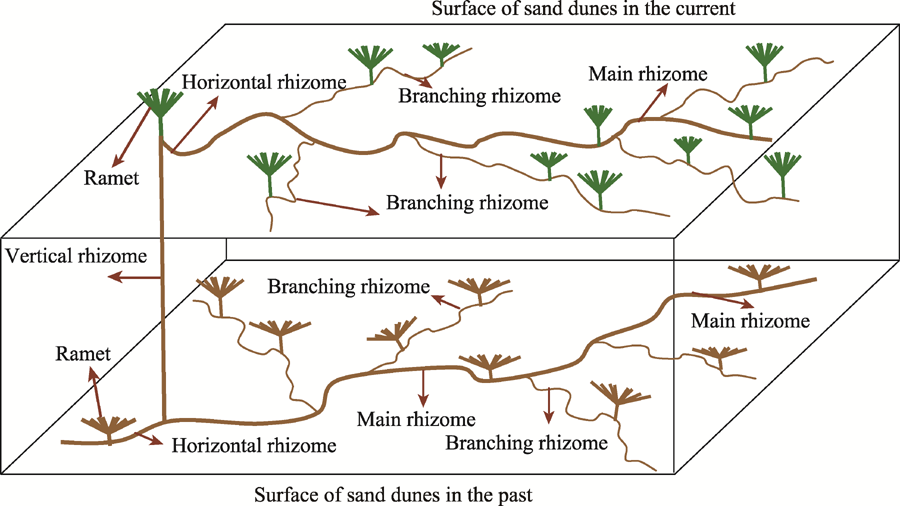
Observations of profile characteristics of C. brunnescen in semi-fixed sand dunes showed that the belowground rhizomes were highly developed and exhibited complex horizontal rhizomes (including branching rhizomes and main rhizomes) and vertical rhizomes (Fig. 2). Ramets were mainly reproduced by the horizontal rhizomes. During the growing season, the ground surface of sand dunes was covered with sands, and the vertical rhizomes that were created by the nodes of the continuously-expanding horizontal rhizomes emerged through the sand surface to form new ramets. The process repeated and then formed a multi-tiered network that stabilized (fixed) the sand dunes. The network of belowground rhizomes formed 4 tiers in a 120-cm sand depth. The maximum sand burial depth from which vertical rhizomes emerged to form new ramets was less than 50 cm.
Fig. 2 Sketch map of the growth and reproductive characteristics of belowground rhizomes of Carex brunnescens
3.2 Morphological characteristics of belowground rhizomes of C. brunnescens
In semi-fixed sand dunes, the average internode length was 3.0 cm with 10 buds on vertical rhizomes (sand burial depth of 50 cm); while in moving sand dunes, the average internode length was 2.5 cm with 10 buds on vertical rhizomes (sand burial depth of 50 cm) (Table 1). For branching rhizomes, the average internode lengths were 2.5 and 2.6 cm in semi-fixed and moving sand dunes, respectively; while for main rhizomes, the average internode lengths were 4.0 and 4.1 cm in semi-fixed and moving sand dunes, respectively. There was a significant difference in the average internode length between branching rhizomes and main rhizomes in both semi-fixed and moving sand dunes (Table 1).
Table 1
Table 1
Table 1 Morphological characteristics of belowground rhizomes of C. brunnescens in semi-fixed and moving sand dunes
Rhizomes
Semi-fixed sand dunes
Moving sand dunes
Length (cm)
Internode length (cm)
Bud number of rhizome nodes
Length (cm)
Internode length (cm)
Bud number of rhizome nodes
Vertical
40-50
3.0± 0.6ab
10± 1
40-50
2.5± 0.3b
9± 2
Branching
Min: 230± 80
2.5± 0.2b
Min: 96± 6
Min: 280± 60
2.6± 0.3b
Min: 108± 7
Max: 1260± 120
Max: 504± 17
Max: 1530± 180
Max: 585± 20
Main
Min: 410± 60
4.0± 0.7a
Min: 98± 7
Min: 630± 90
4.1± 0.5a
Min: 154± 9
Max: 3680± 240
Max: 924± 22
Max: 4520± 360
Max: 1103± 28
Note: Min, minimum; Max, maximum. Different lowercase letters within a column indicate significant differences among different rhizome types at P< 0.05 level (Duncan’ s test). Mean± SD.
Table 1 Morphological characteristics of belowground rhizomes of C. brunnescens in semi-fixed and moving sand dunes
3.3 Relationships of vegetation parameters of C. brunnescens with wind erosion height and sand burial depth
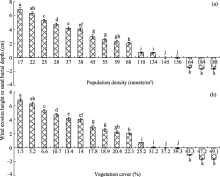
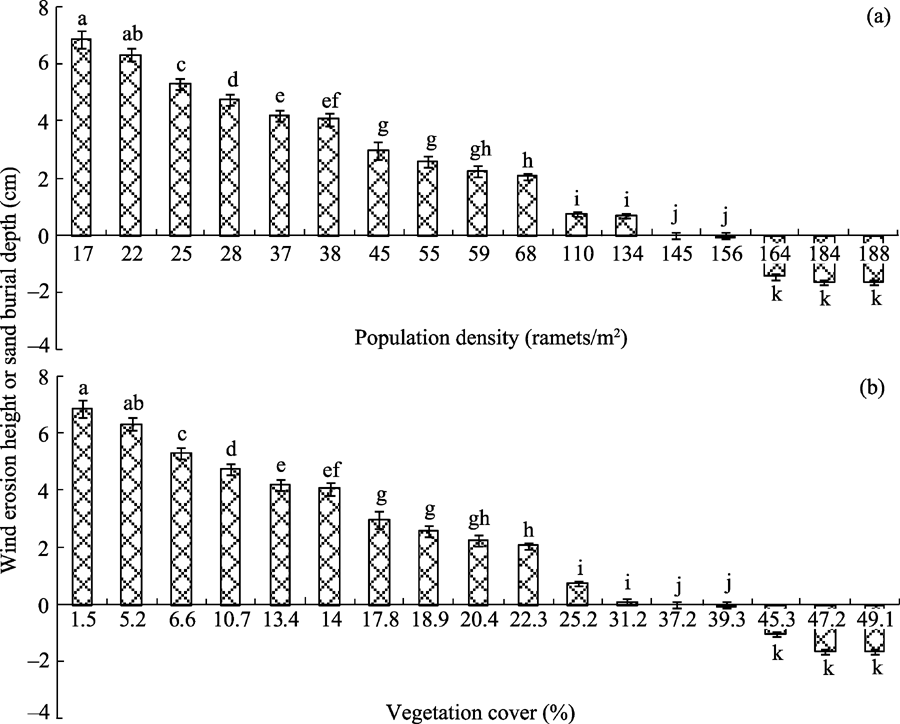
Population density and vegetation cover of C. brunnescens greatly influenced wind erosion height and sand burial depth (Fig. 3). Generally speaking, wind erosion heights decreased with increases in population density (0-145 ramets/m2) and vegetation cover (1.5%-29.2%). It should be mentioned that wind erosion was absent at population density of 145-156 ramets/m2and vegetation cover of 31.2%-39.3%. Furthermore, apparent sand burial was observed at population density > 156 ramets/m2and vegetation cover > 39.3%.
Fig. 3 Wind erosion height or sand burial depth under different (a) population densities and (b) vegetation covers of C. brunnescens. Bars mean standard deviations; n=5. Different lowercase letters indicate significant differences of wind erosion height (or sand burial depth) among different population densities and among different vegetation covers of C. brunnescens at P< 0.05 level (Duncan’ s test). It should be noted that positive value denotes wind erosion height while negative value denotes sand burial depth.
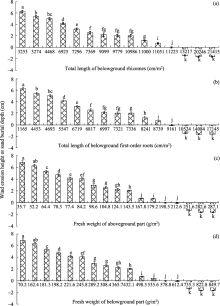
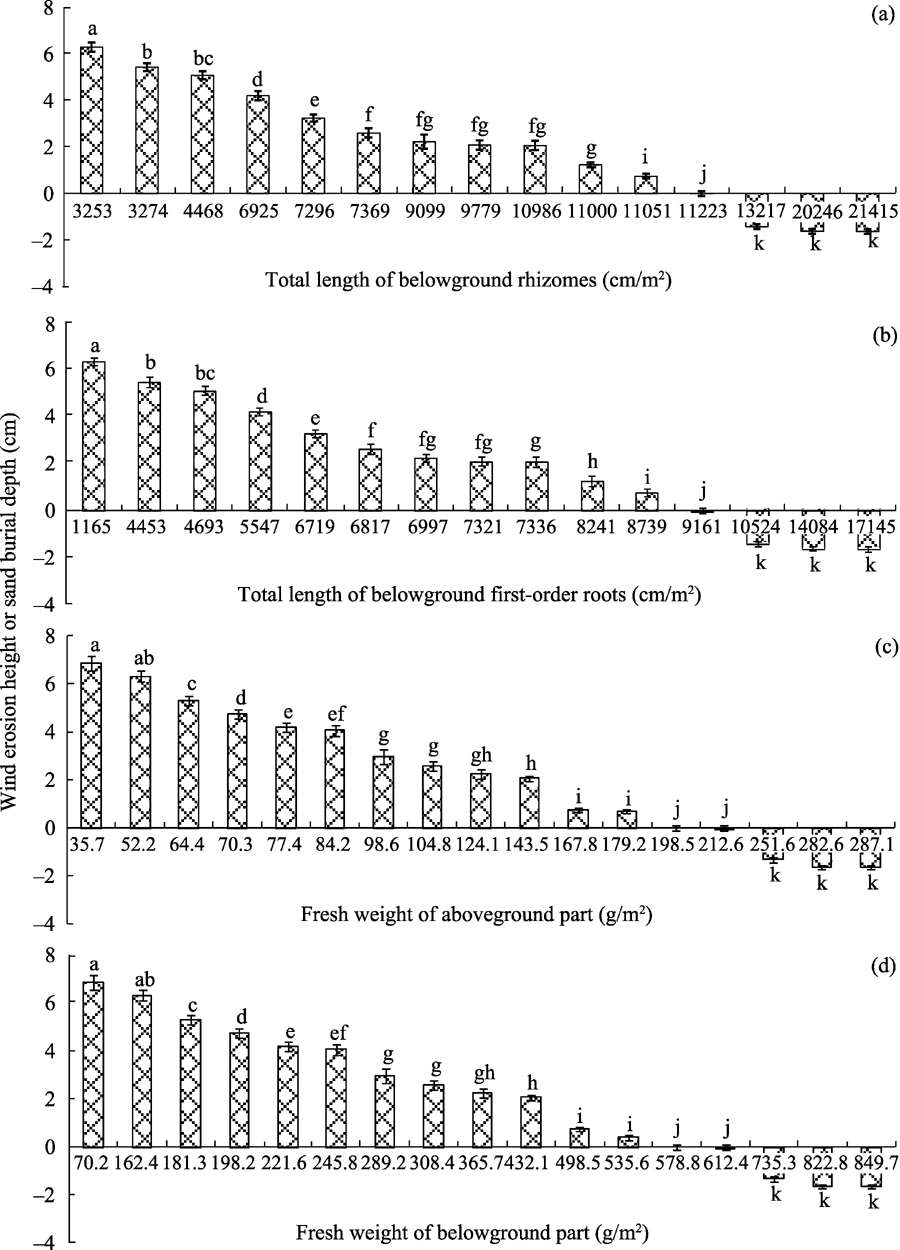
For C. brunnescens, the total length of belowground rhizomes, total length of belowground first-order roots, fresh weight of aboveground part and fresh weight of belowground part exerted great influences on wind erosion height and sand burial depth (Fig. 4). Wind erosion heights decreased with increases in total length of belowground rhizomes (0-11, 051 cm/m2), total length of belowground first-order roots (0-8739 cm/m2), fresh weight of aboveground part (35.7-179.2 g/m2) and fresh weight of belowground part (70.2-535.6 g/m2). It should be mentioned that wind erosion was not observed under following conditions: total length of belowground rhizomes being 11, 223 cm/m2, total length of belowground first-order roots being 9161 cm/m2, fresh weight of aboveground part being 198.5-212.6 g/m2and fresh weight of belowground part being 578.8-612.4 g/m2. Furthermore, apparent sand burial occurred under following conditions: total length of belowground rhizomes > 11, 223 cm/m2, total length of belowground first-order roots > 9161 cm/m2, fresh weight of aboveground part > 212.6 g/m2 and fresh weight of belowground part > 612.4 g/m2.
Fig. 4 Wind erosion height or sand burial depth under different (a) total lengths of belowground rhizomes, (b) total lengths of belowground first-order roots, (c) fresh weight of aboveground part and (d) fresh weight of belowground part of C. brunnescens. Bars indicate standard deviations; n=5. Different lowercase letters indicate significant differences of wind erosion height (or sand burial depth) among different growth-related values of C. brunnescens at P< 0.05 level (Duncan’ s test). It should be noted that positive value denotes wind erosion height while negative value denotes sand burial depth.
3.4 Growth and reproductive performance of C. brunnescens in three experimental sites
As shown in Table 2, applying SCT in sand dunes and transplanting belowground rhizomes of C. brunnescens to sand dunes with SCT significantly improved the growth performance of C. brunnescens. In September 2013, the values of ramet height, total number of reproductive ramets, total number of belowground rhizomes, total length of belowground rhizomes, and fresh weight of C. brunnescens were 39.6%, 323.5%, 64.7%, 69.3%, and 123.1% higher in SD+SCT+P than in SD+N, respectively. And, those values were 21.8%, 141.1%, 28.3%, 18.0%, and 42.8% higher in SD+SCT+P than in SD+SCT, respectively. The effects of applying SCT in sand dunes and transplanting belowground rhizomes to sand dunes with SCT on the growth and reproductive performance of C. brunnescens were also confirmed in September 2014 and 2015.
Table 2
Table 2
Table 2 Growth-related parameters of C. brunnescens in three experimental sites in different periods
Period
Experimental site
RH (cm)
TRR (numbers/m2)
TBR (numbers/m2)
TLR (cm/m2)
FW (g/m2)
SD+N
4.8± 0.3d
13.6± 2.3h
28.6± 2.6f
789.3± 41.4g
204.5± 11.6h
September 2013
SD+SCT
5.5± 0.3c
26.9± 3.3f
36.7± 2.9e
1132.4± 67.4f
319.6± 12.3f
SD+SCT+P
6.7± 0.4ab
57.6± 4.6c
47.1± 3.1d
1336.4± 82.1de
456.3± 29.8de
SD+N
4.6± 04de
23.7± 2.9fg
41.8± 3.4de
1013.6± 69.8cd
275.6± 20.2g
September 2014
SD+SCT
6.3± 0.4b
37.6± 3.7e
48.3± 3.6d
1317.8± 78.2de
463.6± 48.9d
SD+SCT+P
7.1± 0.6a
83.5± 8.1b
69.2± 5.1b
1795.6± 108.4b
694.8± 30.1b
SD+N
4.9± 0.5d
37.2± 2.4e
56.3± 3.1c
1324.5± 76.6d
326.5± 23.2f
September 2015
SD+SCT
6.1± 0.3b
51.4± 3.3cd
72.6± 3.3b
1517.7± 88.2c
668.6± 33.6bc
SD+SCT+P
7.0± 0.6a
116.7± 9.2a
88.4± 4.5a
1976.3± 101.5a
1021.4± 51.2a
Note: RH, ramet height; TRR, total number of reproductive ramets; TBR, total number of belowground rhizomes; TLR, total length of belowground rhizomes; FW, fresh weight. SD+N, sand dunes that had no further treatment; SD+SCT, sand dunes that had straw checkerboard technique but no transplanted rhizomes of C. brunnescens; SD+SCT+P, sand dunes that had both straw checkerboard technique and transplanted rhizomes of C. brunnescens. Experimental sites of SD+N, SD+SCT and SD+SCT+P in Table 3 present the same meanings as this table. Mean± SD; n=5. Different lowercase letters within the same column indicate significant differences among different experimental sites and different periods at P< 0.05 level (Duncan’ s test).
Table 2 Growth-related parameters of C. brunnescens in three experimental sites in different periods
3.5 Effects of belowground rhizomes of C. brunnescen and SCT on soil properties
Applying SCT in sand dunes and transplanting belowground rhizomes of C. brunnescens to sand dunes with SCT had significant effects on soil nutrient properties (Table 3). Soil nutrient contents (0-15 cm soil depth) significantly increased in sand dunes after the application of SCT and establishment of belowground rhizomes of C. brunnescens. Specifically, the contents of SOC, AN, AP, and AK were 50.4%, 78.4%, 63.1%, and 131.9% higher in SD+SCT than in SD+N, respectively. And, those values were 34.5%, 23.5%, 43.0%, and 25.4% higher in SD+SCT+P than in SD+SCT, respectively. Similar variation trends were also observed in 15-30 and 30-50 cm soil depths. Furthermore, SOC, AN, AP, and AK contents exhibited generally decreasing trends with increasing soil depth from April 2012 to September 2015.
Table 3
Table 3
Table 3 Soil nutrient properties at different soil depths in three experimental sites from April 2012 to September 2015
Soil depth (cm)
Experimental site
SOC
TN
TP
AK
AN
AP
(mg/kg)
(μ g/kg)
SD+N
391± 11f
73± 4de
6.7± 0.5d
65.3± 7f
993.2± 50f
296.4± 22e
0-15
SD+SCT
588± 22c
90± 4cd
7.9± 0.4bc
151.4± 6b
1771.4± 93b
483.5± 20bc
SD+SCT+P
791± 18a
149± 6a
10.6± 0.6a
189.8± 11a
2187.8± 110a
691.2± 32a
SD+N
313± 12h
67± 3ef
4.4± 0.4ef
49.6± 6g
799.2± 65gh
221.7± 18f
15-30
SD+SCT
484± 15d
88± 6cd
7.4± 0.6c
110.1± 8cd
1379.4± 121cd
344.6± 18d
SD+SCT+P
661± 13b
134± 7b
8.6± 0.5b
148.5± 8b
1521.8± 103c
511.5± 27b
SD+N
236± 8i
62± 3f
3.6± 0.5g
40.6± 5gh
612.6± 48e
166.9± 11g
30-50
SD+SCT
384± 15fg
81± 5d
5.0± 0.7e
77.8± 5e
877.8± 72g
288.4± 20e
SD+SCT+P
472± 16de
97± 6c
6.5± 0.6d
121.5± 7c
1043.7± 80e
352.6± 16d
Note: SOC, soil organic carbon; TN, total nitrogen; TP, total phosphorus; AK, available potassium; AN, available nitrogen; AP, available phosphorus. Mean± SD; n=5. Different lowercase letters within the same column indicate significant differences among different experimental sites and different soil depths at P< 0.05 level (Duncan’ s test).
Table 3 Soil nutrient properties at different soil depths in three experimental sites from April 2012 to September 2015
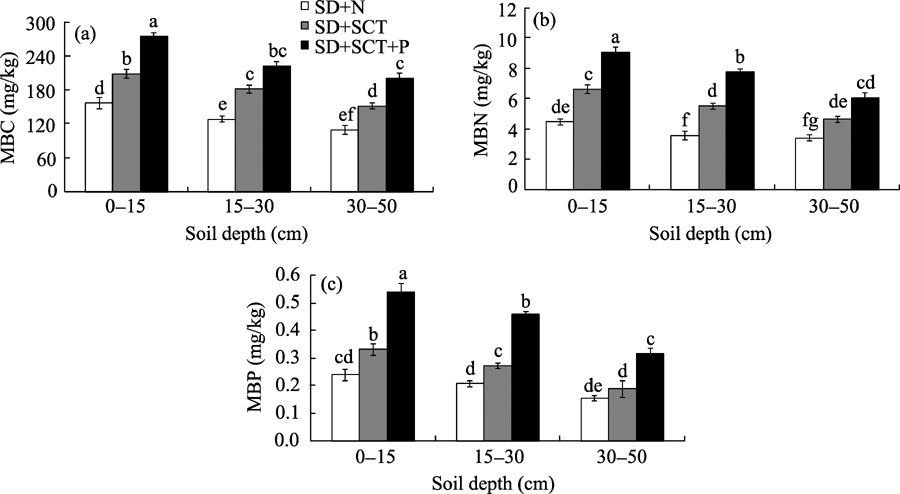
Transplanting belowground rhizomes of C. brunnescens to SD+SCT experimental site significantly affected soil MBC, MBN, and MBP. As shown in Figure 5, from April 2012 to September 2015, the contents of MBC, MBN, and MBP (0-15 cm depth) in SD+SCT+P increased by 33.1%, 48.2%, and 38.3% compared to those in SD+SCT, and by 75.2%, 102.9%, and 125.4% compared to those in SD+N, respectively. Similar variation trends were also found in 15-30 and 30-50 cm soil depths during this period. Furthermore, MBC, MBN, and MBP contents generally decreased with increasing soil depth at each experimental site.
Fig. 5 Soil microbial biomass carbon, nitrogen, and phosphorus at different soil depths in three experimental sites from April 2012 to September 2015. MBC, microbial biomass carbon; MBN, microbial biomass nitrogen; MBP, microbial biomass phosphorus. SD+N, sand dunes that had no further treatment; SD+SCT, sand dunes that had straw checkerboard technique but no transplanted rhizomes of C. brunnescens; SD+SCT+P, sand dunes that had both straw checkerboard technique and transplanted rhizomes of C. brunnescens. Bars indicate standard deviations; n=5. Different lowercase letters indicate significant differences among different experimental sites for the same soil depth at P< 0.05 level (Duncan’ s test).
4 Discussion
4.1 Reproductive characteristics of C. brunnescens and adaptation of C. brunnescens to alpine environments
Clonal plants are special types of plants that can maintain normal survival and reproduction in sand dune environments. During the evolutionary process, clonal plants have acquired many adaptive mechanisms to tolerate sand burial and wind erosion stresses (Liu et al., 2007; Yu et al., 2008; Tang et al., 2010; Ye and Dong, 2011). In this study, we determined that belowground rhizomes of C. brunnescens developed structures with a strong ability to grow upward and to form new ramets when sand burial depth was < 50 cm (see Table 1), indicating that belowground structures of C. brunnescens may be an inevitable evolution under wind stresses (Kang et al., 2016a, b; Ma et al., 2017). Moreover, our results showed that C. brunnescens growing in sand dunes with application of straw checkerboard technique (i.e., SD+SCT) can produce more clonal ramets than that growing in sand dunes that had no further treatment (SD+N), and this trend can be further increased after the establishment of belowground rhizomes of C. brunnescens. This may be due to the positive effects of wind erosion and sand burial on ramet growth and regeneration of C. brunnescens (see Table 2). Interestingly, our previous studies have shown that C. brunnescens generated more ramets in simulated blowouts than in natural conditions and that the sources of new ramets were dormant buds and buds spreading from outside (Kang et al., 2016a). Dormant buds were released from dormancy in the deeper soil layers under wind erosion conditions, suggesting that C. brunnescens has a strong potential for blowout remediation (Kang et al., 2016a). It seems that C. brunnescensshows excellent capacity for sand fixation through rapid clonal expansion that ensures its rapid growth and reproduction. This finding provides a new perspective for improving clonal plants for biomass production and the restoration of degraded vegetation in Maqu alpine region. However, it is still unknown why C. brunnescens grows well in these stressful conditions without any detectable signs of stress from wind erosion or sand burial. The answers may involve division of labor and ramet specialization as important parts of clonal integration in C. brunnescens (Xu et al., 2010; Luo et al., 2015; Luo and Zhao, 2015). Spatial division of labor among ramets that are connected with each other has a strong ability to share resources (including water, carbon, nitrogen, and phosphorus), and ramet specialization and cooperation in resource uptake are beneficial to plant biomass and clonal regeneration of C. brunnescens at small spatial scales (Roiloa et al., 2007, 2014; Luo and Zhao, 2015; Kang et al., 2016a). Therefore, our results further confirmed that clonal integration and reproduction may be extremely important to the survival and regeneration of C. brunnescens in building resistance and resilience to the unstable sand dune environments (Xu et al., 2010; Luo et al., 2015; Kang et al., 2016a). However, the physiological and ecological adaptive mechanisms still need a further study.
4.2 Effects of growth-related characteristics of C. brunnescens on wind erosion and sand burial
Wind erosion and sand burial are the most common abiotic stresses for plant growth in the desert areas of northwestern China. Therefore, the choice of well-adapted plant species for the restoration and stabilization of moving sand dunes is of critical importance (He et al., 2007; Gong et al., 2014; Wang et al., 2016). When vegetation cover decreases, the protective effect of vegetation on surface soil decreases, the exposed part of the surface soil increases, and the small particles of surface soil can be readily blown into the air under the action of strong winds (Gu et al., 2002; Zhang et al., 2016). A significant negative correlation between vegetation cover and wind erosion has been previously documented (Hai et al., 2002; Xu and Liu, 2012; Zhang et al., 2016). Our results showed that forC. brunnescens species, following conditions were well-documented to have great effects on wind erosion and sand burial (see Figs. 3 and 4): population density at 145-156 ramets/m2, vegetation cover at 31.2%-39.3%, total length of belowground rhizomes at 11, 223 cm/m2, total length of belowground first-order roots at 9161-10, 524 cm/m2, fresh weight of aboveground part at 198.5-212.6 g/m2, and fresh weight of belowground part at 578.8-612.4 g/m2. Understanding the vegetation characteristics (composition, structure, and function) of C. brunnescens and addressing the relationships of growth-related characteristics (population density, plant cover, and aboveground and belowground parts) of C. brunnescens with wind erosion and sand burial have important theoretical significance for the prevention and control of soil wind erosion. More importantly, the population and community structure, and configuration and succession of belowground structures of C. brunnescens need to be explored in-depth for promoting natural succession and also for restoring degraded vegetation in Maqu alpine region. To sum up, C. brunnescens can be selected for ecological restoration and soil erosion control in Maqu alpine region.
4.3 Sand fixation with herbs and its application potential with SCT
It is well known that soil erosion is an important process that affects both surface features and biological potential of soils (Wezel et al., 2000; Ye and Dong, 2011; Xu et al., 2012). Sand fixation with plants is considered to be the most effective measure in controlling desertification (Wang et al., 2016). However, vegetation in sand dunes can be easily disturbed by environment changes and human activities. As self-recover ability of vegetation in sand dunes is very weak, a combination of biological and engineering measures is recommended to improve the effect of vegetation on sand stabilization (Dai et al., 2008; Zhang et al., 2014). Previous studies have widely reported that establishment of stable artificial sand-fixing plant communities with SCT is an effective strategy for sand fixation and for soil and water conservation in many arid and semi-arid regions of China (Qiu et al., 2004; Ma et al., 2015). For moving sand dunes, the application of SCT with plants not only stabilizes the surface of sand dunes but also improves soil nutrient properties through belowground root systems and inputs of organic matter from decomposing straw, dead branches, and fallen leaves of plants. It was previously reported that some shrub plants (e.g., Caragana microphyla, Hedysarum fruticosumand Salix oritrepha) and herbs (e.g., Agropyron cristatum) could be planted with SCT to further improve the windbreak and sand dune fixation effects (Qiu et al., 2004; Cao et al., 2007; Zhao et al., 2010; Zhu et al., 2012; Kang et al., 2015). In our study, transplanting belowground rhizomes ofC. brunnescens to SD+SCT not only significantly promoted the growth and reproductive performance of C. brunnescens (Tables 2 and 3) but also increased soil nutrient contents. Our results indicated that the establishment of straw checkerboard in sand dunes increases soil MBC, MBN, and MBP contents, and these contents can be further increased by establishing belowground rhizomes ofC. brunnescens (see Fig. 5). This may be associated with the organic and inorganic substances of plant matter in the soil that provide nutrition and energy for a variety of microorganisms and are conducive to the growth and reproduction of microorganisms.
5 Conclusions
Understanding the vegetation characteristics (composition, structure, and function) of C. brunnescens and addressing the relationships of growth-related characteristics (population density, plant cover, and aboveground and belowground parts) of C. brunnescens with wind erosion and sand burial have important theoretical significance for the prevention and control of soil wind erosion. The efficient vegetative reproduction and high productivity of C. brunnescens in alpine sandy environment are resulted from the highly-developed and complex belowground horizontal rhizomes that can continuously create vertical rhizomes emerging from sand surface and forming new ramets. Our observations demonstrated that transplanting belowground rhizomes of C. brunnescens to SCT-controlled sand dunes has pronounced windbreak and sand fixation effects in degraded alpine sandy environments. Our results further confirmed that clonal integration and reproduction may be extremely important to the survival and regeneration of C. brunnescens in building resistance and resilience to the unstable sand dune environments. However, the physiological and ecological adaptive mechanisms still need a further study. It is also worth mentioning that due to the high seed dormancy of C. brunnescens in natural conditions (seed germination rate < 14%), the practice of using seed reproduction may limit the ability to cultivate C. brunnescens under SCT-controlled sand dunes in the degraded alpine meadow of Maqu. Therefore, the sand fixing principle, methods for breaking seed dormancy, seed germination, and dormancy mechanisms of C. brunnescens also require a further research.
Acknowledgements
This work was supported by the National Natural Science Foundation of China (31360087, 31360086). The authors are very grateful to Dr. Kathryn PIATEK (a professional translator and editor in USA), as well as the anonymous reviewers and editors for their critical review and comments which helped to improve and clarify the paper.
The authors have declared that no competing interests exist.
AmsberryL, Baker MA, Ewanchuk PJ, et al. 2000. Clonal integration and the expansion of Phragmites australis. Ecological Applications, 10(4): 1110-1118. [Cited Within:1]
[2]
Bao SD. 2000. Soil and Agricultural Chemistry Analysis (3rd ed. ). Beijing: China Agriculture Press, 39-86. (in Chinese)[Cited Within:2]
[3]
Brookes PC, Powlson DS, Jenkinson DS. 1982. Measurement of microbial biomass phosphorus in soil. Soil Biology and Biochemistry, 14(4): 319-329. [Cited Within:1]
[4]
Cailson TN, Ripley DA. 1997. On the relation between NDVI, fractional vegetation cover, and leaf area index. Remote Sensing of Environment, 62(3): 241-252. [Cited Within:1]
[5]
Cao YC, Jiang DM, Zhu LH, et al. 2007. A melioration effect of sand -fixing Hedysarum fruticosum plantations on soil nutrient contents and biological activities. Chinese Journal of Applied Ecology, 18(8): 1739-1744. (in Chinese)[Cited Within:1]
[6]
Chen ZC, LiN, Liu CH. 2013. Effects of straw checkerboard barriers on soil nutrients in water diversion project in the Gurbantunggut Desert. Pratacultural Science, 30(5): 699-702. (in Chinese)[Cited Within:1]
[7]
Dai HL, Jin FX, Zhang KL. 2011. A review of field measurement on wind erosion. Advances in Earth Science, 26(4): 401-408. (in Chinese)[Cited Within:1]
[8]
Dai SM, Qiu GY, ZhaoM. 2008. Study on land desertification and its prevention and control measures in the Minqin Oasis in Gansu Province. Arid Zone Research, 25(3): 319-324. (in Chinese)[Cited Within:1]
[9]
Dech JP, Maun MA. 2006. Adventitious root production and plastic resource allocation to biomass determine burial tolerance in woody plants from central Canadian coastal dunes. Annals of Botany, 98(5): 1095-1105. [Cited Within:1]
[10]
Diao ZY, Xu LR, Feng CY, et al. 2012. The ground spectral model for estimating vegetation coverage on desertified grassland , Hulunbeier, Inner Mongolia, China. Journal of Arid Land Resources and Environment, 26(2): 139-144. (in Chinese)[Cited Within:1]
[11]
Dong ZB, Wang XM, Liu LY. 2000. Wind erosion in arid and semiarid China: An overview. Journal of Soil and Water Conservation, 55(4): 439-444. [Cited Within:1]
[12]
GomesL, Arrúe JL, López MV, et al. 2003. Wind erosion in a semiarid agricultural area of Spain: The WELSONS project. CATENA, 52(3-4): 235-256. [Cited Within:1]
[13]
Gong GL, Liu JY, Shao QQ. 2014. Effects of vegetation coverage change on soil conservation service of typical steppe in Inner Mongolia. Journal of Geo-informationScience, 16(3): 426-434. (in Chinese)[Cited Within:1]
[14]
GuW, Cai XP, XieF, et al. 2002. Study on relationship between vegetation cover and distribution of days of sand storm: Taking central and western inner mongolia for example. Advance in Earth Sciences, 17(2): 273-277. (in Chinese)[Cited Within:1]
[15]
Hai CX, Liu BY, ZhaoY. 2002. Influence of soil humidity and vegetation coverage on wind erosion. Chinese Journal of Applied Ecology, 13(8): 1057-1058. (in Chinese)[Cited Within:1]
[16]
He SF, Jiang DM, Alamusa. 2007. Roles of vegetation in wind erosion control: A research review. Chinese Journal of Ecology, 26(5): 743-748. (in Chinese)[Cited Within:1]
[17]
Jiang DM, Zhou QL, Li XH, et al. 2008. Sand -stabilizing measures and effects of biology and engineering of shifting sand in Horqin sand y land . Journal of Liaoning Technical University (Natural Science), 27(1): 141-143. (in Chinese)[Cited Within:2]
[18]
Jiang SL, Hu YF, PuQ, et al. 2016. Changes in soil nitrogen characteristics during grassland desertification in Northwest Sichuan. Acta Ecologica Sinica, 36(15): 4644-4653. (in Chinese)[Cited Within:1]
[19]
Kang JJ, Duan JJ, Wang SM, et al. 2013. Na compound fertilizer promotes growth and enhances drought resistance of the succulent xerophyte Haloxylon ammodendron. Soil Science and Plant Nutrition, 59(2): 289-299. [Cited Within:2]
[20]
Kang JJ, Zhao WZ, ZhaoM, et al. 2015. The cutting reproduction technique of Salix oritrepha and its application on degraded grassland s restoration. Journal of Soil and Water Conservation, 70(1): 45-53. [Cited Within:7]
[21]
Kang JJ, Zhao WZ, ZhaoM. 2016a. Remediation of blowouts by clonal plants in Maqu degraded alpine grassland s of northwest China. Journal of Plant Research, 31(2): 291-299. [Cited Within:9]
[22]
Kang JJ, ZhaoM, Li GY, et al. 2016b. Tentative research of sand -fixing characteristics of Carex brunnescens on desertified grassland in Maqu alpine regions. Soil and Water Conservation in China, (2): 47-51. (in Chinese)[Cited Within:5]
[23]
Li SL, Werger M JA, Zuidema PA, et al. 2010a. Seedlings of the semi-shrub Artemisia ordosica are resistant to moderate wind denudation and sand burial in Mu Us sand land , China. Trees, 24(3): 515-521. [Cited Within:2]
[24]
Li SL, Zuidema PA, Yu FH, et al. 2010b. Effects of denudation and burial on growth and reproduction of Artemisia ordosica in Mu US sand land . Ecological Research, 25(3): 655-661. [Cited Within:1]
[25]
LianJ, Zhao XY, Wang SK, et al. 2013. Impacts of wind erosion on the distribution patterns of soil organic carbon and total nitrogen in Horqin Sand y Land , China. Chinese Journal of Ecology, 32(3): 529-535. (in Chinese)[Cited Within:1]
Luo WC, Zhao WZ, Zeng FJ, et al. 2015. Water but not photosynthates integration exists between mother and daughter ramets of a root-derived clonal shrub. Plant Ecology, 216(2): 331-342[Cited Within:4]
[29]
MaH, Kang JJ, ZhaoM, et al. 2017. Biological sand -fixation characteristics of Carex brunnescens in Yellow River source region. Pratacultural Science, 34(4): 714-723. [Cited Within:3]
[30]
Ma XX, Wang HF, Li SY, et al. 2015. Comparison of sand -protecting efficiency and terrain adapted of two checkerboard barriers. Bulletin of Soil and Water Conservation, 35(2): 344-349. (in Chinese)[Cited Within:3]
[31]
Meadows ME, Hoffman MT. 2002. The nature, extent and causes of land degradation in South Africa: Legacy of the past, lessons for the future?Area, 34(4): 428-437. [Cited Within:1]
[32]
Qi DC, Li GY, Chen WY, et al. 2006. Present status, causes and control countermeasures of natural grassland degeneration in Maqu County. Journal of Desert Research, 26(2): 202-207. (in Chinese)[Cited Within:1]
[33]
Qiu GY, Lee IB, ShimizuH, et al. 2004. Principles of sand dune fixation with straw checkerboard technology and its effects on the environment. Journal of Arid Environments, 56(3): 449-464. [Cited Within:5]
[34]
Roiloa SR, AlpertP, TharayilN, et al. 2007. Greater capacity for division of labour in clones of Fragaria chiloensis from patchier habitats. Journal of Ecology, 95(3): 397-405. [Cited Within:1]
[35]
Roiloa SR, AnteloB, RetuertoR. 2014. Physiological integration modifies δ15N in the clonal plant Fragaria vesca, suggesting preferential transport of nitrogen to water-stressed offspring. Annals of Botany, 114(2): 399-411. [Cited Within:2]
[36]
SDRS (Shapotou Desert Research Station, Institute of Desert Research, Academia Sinica, Lanzhou). 1986. The principles and measures taken to stabilize shifting sand s along the railway line in the southeastern edge of the Tenggar Desert. Journal of Desert Research, 6(3): 1-19. (in Chinese)[Cited Within:1]
[37]
Tang JB, XiaoY, An SQ. 2010. Advance of studies on rhizomatous clonal plants ecology. Acta Ecologica Sinica, 30(11): 3028-3036. (in Chinese)[Cited Within:1]
[38]
WangF, ZhuL, ZhaoM, et al. 2013. Effect of different treatments on seed germination of Carex brunnescens. Chinese Agricultural Science Bulletin, 29(22): 36-39. (in Chinese)[Cited Within:1]
[39]
WangG, QingJ, ChengG, et al. 2001. Eco-environmental degradation and causal analysis in the source region of the Yellow River. Environmental Geology, 40(7): 884-890. [Cited Within:1]
[40]
Wang JB, Wang ZG, LvH. 2008. Climate background analysis of grassland degradation in the important water source supply area of the Yellon River: a case study of Maqu County. Pratacultural Science, 25(4): 23-27. (in Chinese)[Cited Within:1]
[41]
Wang YB, Wang GX, Shen YP, et al. 2005. Degradation of the eco-environmental system in alpine meadow on the Tibetan Plateau. Journal of Glaciology and Geocryology, 27(5): 633-640. (in Chinese)[Cited Within:1]
[42]
Wang YW, ChaiQ, Ouyang XZ, et al. 2016. Evolution of soil nutrition and biological prosperities under sand -fixing plants in fringe area of Minqin oasis. Journal of Soil and Water Conservation, 30(1): 141-146, 177. (in Chinese)[Cited Within:2]
[43]
WeiQ, WangF, Chen WY, et al. 2010. Soil physical characteristics on different degraded alpine grassland s in Maqu County in upper Yellow River. Bulletin of Soil and Water Conservation, 30(5): 16-21. (in Chinese)[Cited Within:1]
[44]
WezelA, Rajot JL, HerbrigC. 2000. Influence of shrubs on soil characteristics and their function in Sahelian agro-ecosystems in semi-arid Niger. Journal of Arid Environments, 44(4): 383-398. [Cited Within:1]
[45]
Xing XY, Huang YM, An SS, et al. 2013. Soil nitrogen concentrations and transformations under different vegetation types in forested zones of the Loess Gully region. Acta Ecologica Sinica, 33(22): 7181-7189. (in Chinese)[Cited Within:1]
[46]
Xu CY, Schooler SS, Van Klinken RD. 2010. Effects of clonal integration and light availability on the growth and physiology of two invasive herbs. Journal of Ecology, 98(4): 833-844. [Cited Within:3]
[47]
Xu MH, LiuT. 2012. Species composition and windproof effect of natural vegetation at an oasis-desert ecotone in early spring. Arid Zone Research, 29(1): 64-72. (in Chinese)[Cited Within:3]
[48]
Xu MX, WangZ, ZhangJ, et al. 2012. Response of soil organic carbon sequestration to the “Grain for Green Project” in the hilly Loess Plateau region. Acta Ecologica Sinica, 32(17): 5405-5415. (in Chinese)[Cited Within:1]
[49]
Ye XH, DongM. 2011. Remediation of blowout pits by clonal plants in Mu Us Sand land . Acta Ecologica Sinica, 31(19): 5505-5511. (in Chinese)[Cited Within:3]
[50]
Yu FH, WangN, He WM, et al. 2008. Adaptation of rhizome connections in dryland s: Increasing tolerance of clones to wind erosion. Annals of Botany, 102(4): 571-577. [Cited Within:2]
[51]
Zhang DS, Wu WY, Tian LH, et al. 2014. Effects of erosion and deposition and dimensions selection of straw-checkerboard barriers in the desert of Qinghai Lake. Scientia Geographica Sinica, 34(5): 627-634. [Cited Within:1]
[52]
ZhangP, Yin DZ, Shang HY. 2016. Anti-wind erosion effect of plants in Kara Bailey project area. Bulletin of Soil and Water Conservation, 36(1): 224-229. (in Chinese)[Cited Within:3]
[53]
Zhao HL, He YH, Zhou RL, et al. 2009. Effects of desertification on soil organic C and N content in sand y farmland and grassland of Inner Mongolia. Catena, 77(3): 187-191. [Cited Within:1]
[54]
Zhao WZ, Li QY, Fang HY. 2007. Effects of sand burial disturbance on seedling growth of Nitraria sphaerocarpa. Plant and Soil, 295(1-2): 95-102. [Cited Within:1]
[55]
Zhao XB, DangB, Fu YR, et al. 2010. Technology of wind-breaking and sand -fixing forest establishment along expressway in semiarid sand y land and benefit evaluation. Journal of Desert Research, 30(6): 1247-1255. (in Chinese)[Cited Within:1]
[56]
ZhuL, ZhaoM, Li GY, et al. 2012. Types of secondary barren in Maqu County on the upper Yellow River and its remediation techniques. Soil and Water Conservation in China, (10): 50-52. (in Chinese)[Cited Within:1]
[57]
ZhuL, WangF, ZhaoM, et al. 2013. Study on seed biological characteristics of Carex brunnescens. Soil and Water Conservation in China, (9): 56-58. (in Chinese)[Cited Within:1]
1
2000
0.0
0.0
... Clonal integration is considered to be one of the most important strategies for the survival of clonal plants because it allows buried ramets to access to water and nutrients via non-buried ramets, thus ensuring plant survival and reproduction in the environments that are prone to sand burial and wind erosion (Amsberry et al ...
2
0.0
0.0
... , 2015), total nitrogen (TN) was measured following the Kjeldahl digestion and total phosphorus (TP) was determined by a UV-1601 visible spectrophotometer (Bao, 2000 ...
... In addition, soil available nitrogen (AN) was determined by the alkaline diffusion method, available phosphorus (AP) was determined by the Bray method, and available potassium (AK) was measured using the ammonium acetate extract method (Bao, 2000) ...
1
1982
0.0
0.0
... 5 mm/L K2SO4 extract, microbial biomass nitrogen (MBN) was determined using the Kjeldahl procedure, and microbial biomass phosphorus (MBP) was measured with the NaHCO3 extraction method after soil fumigation with chloroform (Brookes et al ...
1
1997
0.0
0.0
... %) was calculated using the digital imagery (Cailson and Ripley, 1997) ...
1
2007
0.0
0.0
... Cao et al ...
1
2013
0.0
0.0
... Chen et al ...
1
2011
0.0
0.0
... Wind erosion height and sand burial depth were determined for every wind event in the windy season (April and May) by placing soil sample of known mass in a pan-shaped container (wind erosion pan) and later measured by reweighing the samples (Dai et al ...
1
2008
0.0
0.0
... As self-recover ability of vegetation in sand dunes is very weak, a combination of biological and engineering measures is recommended to improve the effect of vegetation on sand stabilization (Dai et al ...
1
2006
0.0
0.0
... Dech and Maun, 2006 ...
1
2012
0.0
0.0
... 0 software to reduce the errors caused by image edge distortion (Diao et al ...
1
2000
0.0
0.0
... However, stimulating effects on plant growth were also observed when plants were subjected to relatively shallow sand burial (Dong et al ...
1
2003
0.0
0.0
... In particular, desertification caused by wind erosion poses a severe threat to agricultural productivity and ecological environment (Gomes et al ...
1
2014
0.0
0.0
... Gong et al ...
1
2002
0.0
0.0
... When vegetation cover decreases, the protective effect of vegetation on surface soil decreases, the exposed part of the surface soil increases, and the small particles of surface soil can be readily blown into the air under the action of strong winds (Gu et al ...
1
2002
0.0
0.0
... A significant negative correlation between vegetation cover and wind erosion has been previously documented (Hai et al ...
1
2007
0.0
0.0
... Therefore, the choice of well-adapted plant species for the restoration and stabilization of moving sand dunes is of critical importance (He et al ...
2
2008
0.0
0.0
... It not only stabilizes the surface of sand dunes but also improves soil qualities through organic inputs of decomposed straw, dead branches and fallen leaves (Jiang et al ...
... Jiang et al ...
1
2016
0.0
0.0
... Jiang et al ...
2
2013
0.0
0.0
... Kang et al ...
... The sampling methods were referred to those described by Kang et al ...
7
2015
0.0
0.0
... because of its vital function in conserving soil and water in the upper reaches of the Yellow River (Kang et al ...
... Kang et al ...
... , 2013, 2015) ...
... 4 mm (Kang et al ...
... Soil organic carbon (SOC) was measured following the K2Cr2O7-H2SO4 oxidation method of Walkley and Black (Kang et al ...
... Kang et al ...
... Kang et al ...
9
2016
0.0
0.0
... Kang et al ...
... ) is considered to be the only herb species found to date that can develop on sand dunes in Maqu alpine region (Kang et al ...
... brunnescens is highly tolerant to wind erosion and also to sand burial because of its rapid growth and regeneration capacities of rhizomes, basal meristem and belowground storage organs (Kang et al ...
... (Kang et al ...
... brunnescens may be an inevitable evolution under wind stresses (Kang et al ...
... brunnescens generated more ramets in simulated blowouts than in natural conditions and that the sources of new ramets were dormant buds and buds spreading from outside (Kang et al ...
... brunnescens has a strong potential for blowout remediation (Kang et al ...
... Kang et al ...
... Kang et al ...
5
2016
0.0
0.0
... , 2016a, b ...
... , 2016a, b) ...
... , 2016a, b ...
... (Kang et al ...
... , 2016a, b ...
2
2010
0.0
0.0
... Sand burial and wind erosion are two of the most serious abiotic stresses on plant growth in many arid and semi-arid areas (Li et al ...
... Li et al ...
1
2010
0.0
0.0
... , 2010a, b) ...
1
2013
0.0
0.0
... Lian et al ...
1
2007
0.0
0.0
... During the evolutionary process, clonal plants have acquired many adaptive mechanisms to tolerate sand burial and wind erosion stresses (Liu et al ...
3
2015
0.0
0.0
... Luo et al ...
... Luo et al ...
... Luo et al ...
4
2015
0.0
0.0
... Luo and Zhao, 2015) ...
... Luo and Zhao, 2015) ...
... Luo and Zhao, 2015) ...
... Luo and Zhao, 2015 ...
3
2017
0.0
0.0
... Ma et al ...
... Ma et al ...
... Ma et al ...
3
2015
0.0
0.0
... Ma et al ...
... Ma et al ...
... Ma et al ...
1
2002
0.0
0.0
... Even worse, regional political instability and social insecurity have also been linked to desertification (Meadows and Hoffman, 2002) ...
1
2006
0.0
0.0
... However, long-term over-exploitation of alpine meadow associated with rapid population increases, environmentally-unfriendly means of agricultural production, overgrazing by livestock, mining, and urbanization inevitably triggered severe desertification in this region (Qi et al ...
5
2004
0.0
0.0
... The straw checkerboard technique (SCT) was devised at the Shapotou Desert Research Station, Chinese Academy of Sciences in 1957 (SDRS, 1986) and has been widely used since for stabilizing sand dunes in arid and semi-arid regions of China (Qiu et al ...
... can be planted to further improve the windbreak and sand dune fixation effects (Qiu et al ...
... 1 m checkerboard pattern, and this pattern was reported to be able to achieve remarkable windbreak and sand dune fixation effects (Qiu et al ...
... Previous studies have widely reported that establishment of stable artificial sand-fixing plant communities with SCT is an effective strategy for sand fixation and for soil and water conservation in many arid and semi-arid regions of China (Qiu et al ...
... , Agropyron cristatum) could be planted with SCT to further improve the windbreak and sand dune fixation effects (Qiu et al ...
1
2007
0.0
0.0
... brunnescens at small spatial scales (Roiloa et al ...
2
2014
0.0
0.0
... Clonal plants, the dominant plant type growing in sand dune environments, have developed multiple adaptive strategies to resist sand burial and wind erosion (Roiloa et al ...
... , 2007, 2014 ...
1
1986
0.0
0.0
... The straw checkerboard technique (SCT) was devised at the Shapotou Desert Research Station, Chinese Academy of Sciences in 1957 (SDRS, 1986) and has been widely used since for stabilizing sand dunes in arid and semi-arid regions of China (Qiu et al ...
1
2010
0.0
0.0
... Tang et al ...
1
2013
0.0
0.0
... brunnescens rarely reproduces sexually even in high-precipitation months or in wet years (Wang et al ...
1
2001
0.0
0.0
... The situation has been worsened by human alteration of river channels and also by human withdrawal of groundwater in the Yellow River (Wang et al ...
1
2008
0.0
0.0
... Consequently, the Maqu alpine region has risk to become a main source of sandstorms in China if effective control measures are not implemented (Wang et al ...
1
2005
0.0
0.0
... , 2001, 2005) ...
2
2016
0.0
0.0
... Wang et al ...
... Sand fixation with plants is considered to be the most effective measure in controlling desertification (Wang et al ...
1
2010
0.0
0.0
... Wei et al ...
1
2000
0.0
0.0
... 3 Sand fixation with herbs and its application potential with SCTIt is well known that soil erosion is an important process that affects both surface features and biological potential of soils (Wezel et al ...
1
2013
0.0
0.0
... Xing et al ...
3
2010
0.0
0.0
... Xu et al ...
... brunnescens (Xu et al ...
... brunnescens in building resistance and resilience to the unstable sand dune environments (Xu et al ...
3
2012
0.0
0.0
... 15%, respectively (Xu and Liu, 2012 ...
... Xu and Liu, 2012 ...
... Xu et al ...
1
2012
0.0
0.0
3
2011
0.0
0.0
... Relative to parent plants, the new ramets could achieve clonal expansion horizontally and thus enter into different habitats (Ye and Dong, 2011 ...
... Ye and Dong, 2011) ...
... Ye and Dong, 2011 ...
2
2008
0.0
0.0
... Wind erosion can render soil infertile through the removal of nutrient-rich surface soil and a progressive decrease in vegetation cover (Yu et al ...
... Yu et al ...
1
2014
0.0
0.0
... Zhang et al ...
3
2016
0.0
0.0
... Zhang et al ...
... Zhang et al ...
... Zhang et al ...
1
2009
0.0
0.0
... Desertification involves severe vegetation degradation and intensified wind and water erosions (Zhao et al ...
1
2007
0.0
0.0
... Zhao et al ...
1
2010
0.0
0.0
... Zhao et al ...
1
2012
0.0
0.0
... Zhu et al ...
1
2013
0.0
0.0
... Zhu et al ...
Sand-fixing characteristics of Carex brunnescens and its application with straw checkerboard technique in restoration of degraded alpine meadows




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
 , TAN Yanrong
, TAN Yanrong